
世界最大級の花、ラフレシアが「寄生植物」だということは、知っていますか?しかも、この植物には驚愕の秘密があります。
以前の記事で、光合成をやめた植物の二つの大きな生き方の1つとして、「寄生植物」を紹介しました。今回は、この寄生植物について、意外と知られていないその生態や、国内外の多様な寄生植物を見ていきましょう。

寄生植物にはなんと、宿主から驚きの物質までもらってしまう植物も!!

「寄生植物」とは何か? ― 宿主から直接奪う生き方

出典:WIKIMEDIA COMMONS – CassythaFil9
私たちが「植物」と聞いて思い浮かべるのは、太陽の光を浴び、緑の葉で自ら栄養を生み出す姿かもしれません。しかし植物の世界は、私たちの想像以上に多様です。
中には光合成という基本的な生存手段を部分的あるいは完全に放棄し、他の生きた植物に頼って生きるものたちがいます。それが「寄生植物」です。

一体、どのような仕組みで他の植物から養分を得ているのか、また、この生き方はどのような進化の必然性から生まれたのか、その謎を探っていきましょう!
寄生植物の定義と「吸器」という武器
寄生植物とは、他の生きた植物(「宿主」と呼ばれます)の組織に直接侵入し、そこから水分や養分を奪い取って生きる植物たちの総称です。この定義を実現する最も重要な構造が、「吸器(きゅうき)」と呼ばれる特殊な器官です。
吸器は、寄生植物が宿主に侵入するための多機能な侵攻器官です。根や茎などが変化して形成されたこの器官は、宿主の細胞壁を溶かす酵素を分泌しながら組織内部に徐々に侵入し、最終的には宿主の維管束※に直接接続されます。
この接続が成功すれば、寄生植物は宿主の生命維持システムから直接、必要な資源を引き出すことができるのです。
維管束(いかんそく)🌿💧
維管束(いかんそく)は、植物の根や茎、葉に通っている、水や栄養分を運ぶための「パイプ」のような組織です。維管束は「道管(どうかん)」と「師管(しかん)」という二つの細い管からなっており、道管は根から吸い上げた水と土の中の栄養を上へ運び、師管は葉で作られた栄養分を下へ運びます。
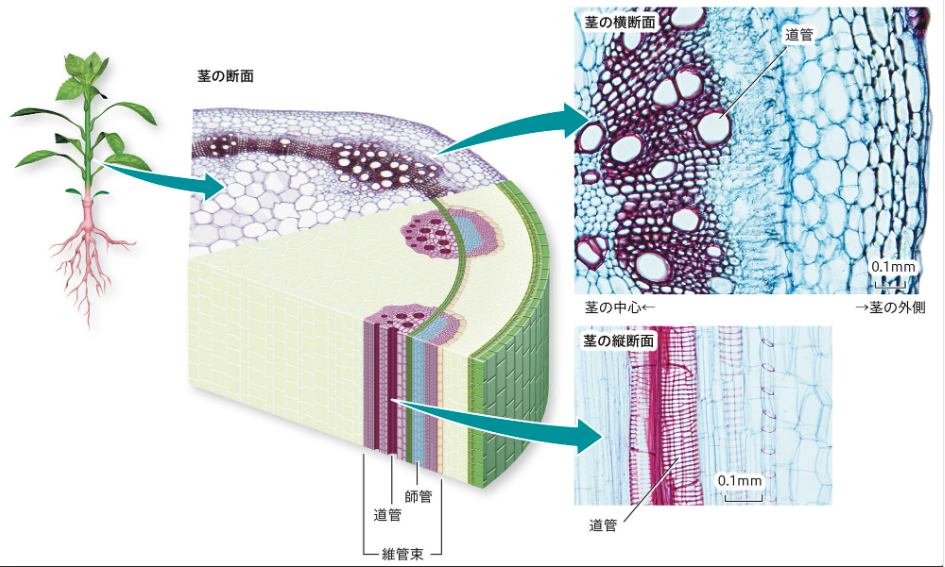
出典:学校図書 ‐ 道管と師管
これら多数の道管と師管が集まった部分を、それぞれ「木部(もくぶ)」「師部(しぶ)」と呼びます。茎の断面を観察すると、維管束が小さな穴(道管)として見えることがあり、茎を太らせるために木部と師部の間には「形成層(けいせいそう)」という細胞分裂の活発な層が存在します。
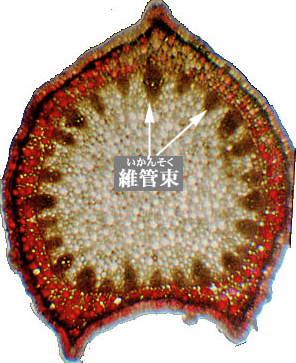
出典:神戸大学 ‐ 専門的な言葉の詳しい解説 維管束 いかんそく
また、葉では維管束が筋状に走っており、これが「葉脈(ようみゃく)」として目に見える構造です。植物全体に水と栄養を効率よく届けるため、欠かせない組織です。

出典:WIKIMEDIA COMMONS – Starr 041211-1365 Unknown pteridophyte
出典:理化学研究所 ‐ 寄生植物の侵入器官発生メカニズムの一端を解明(2016年7月)
吸器の発達は極めて精密なプロセスです。寄生植物が宿主の近くで発芽すると、宿主の根から分泌される化学物質を感知し、この化学シグナルが吸器発達のきっかけとなります。
その後、細胞壁分解酵素(β-1,4-グルカナーゼなど)を宿主に向かって分泌し、宿主の細胞壁を少しずつ溶かしながら侵入していきます。このメカニズムは、人間が行う接ぎ木※の原理と共通する部分があり、植物の組織接合能力がいかに精密であるかを示しています。
接ぎ木🔧 🌱 🔗
接ぎ木は、ある植物の枝(穂木:ほぎ)を、別の植物(台木:だいぎ)につなぎ合わせ、一つの植物として成長させる農業技術です。病気に強い台木の根と、美味しい実がなる穂木の能力を併せ持つ、優れた植物を作ることができます。

出典:WIKIMEDIA COMMONS – Grafting of Dekopon
※左は接ぎ木後に新たな葉が成長、右上は接ぎ木に失敗し枝が枯れる
科学的には、切断面どうしが触れると、植物が持つ「傷を治す力(癒合能力)」が働きます。「カルス」という細胞のかたまりが作られ、お互いの維管束(水や養分の通り道)を接続させるのです。
この組織がくっつく過程で、細胞壁を再構成するために「β-1,4-グルカナーゼ」という酵素が働きます。驚くべきことに、これは寄生植物が吸器で宿主に侵入する際に使う酵素と共通しています。
最新の研究では、植物がこの「つながる能力」を本来持っており、遠く離れた仲間とも接ぎ木できる可能性が示されています。寄生植物は、植物が持つこの基本的な能力を、一方的な「侵略」のために転用していると考えられるのです。
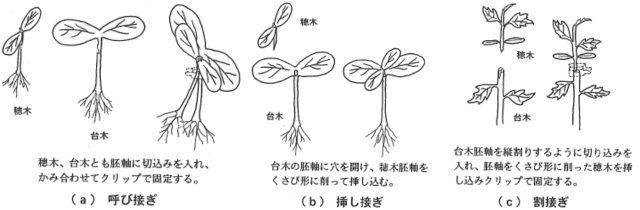
出典:農研機構 ‐ 農業機械化研究所報告書 第31号(1998年3月)
KEN – The mechanism that enables plant grafting is revealed
Science – Cell-cell adhesion in plant grafting is facilitated by β-1,4-glucanases
Phys.org – Cell wall degrading enzyme is integral for plant parasitism and cross-species grafting
- 根寄生植物:地下で宿主の根に寄生(ストライガ、ナンバンキセルなど)
- 茎寄生植物:地上で宿主の茎に寄生(ヤドリギ、ネナシカズラなど)
寄生部位の違いは、宿主認識から侵入、栄養吸収に至るまで、すべてのプロセスに影響を与えます。
全寄生植物と半寄生植物
寄生植物を分類するもう一つの重要な軸が、光合成能力を保持しているかどうかという点です。
全寄生植物
全寄生植物(ぜんきせいしょくぶつ)は、葉緑素を完全に失い、光合成をまったく行いません。そのため、水分と養分のすべてを、生きるために必要な有機栄養(炭水化物など)まで含めて、宿主から奪わなければ生きられません。
ラフレシア 、 ネナシカズラ、 ハマウツボ などがこの分類に該当します。全寄生植物の多くは緑色ではなく、黄色、赤、紫、茶色といった独特の色合いをしており、見た目からも光合成能力を失っていることが推察できます。
半寄生植物
一方、半寄生植物(はんきせいしょくぶつ)は光合成能力を保持しており、自ら有機物を生産することもできます。しかし、それでもなお宿主に寄生しています。
半寄生植物が主に吸収するのは水分や無機栄養(窒素、リンなど)です。 ヤドリギ やコシオガマなどがこれに該当し、特にクリスマスの装飾に使われるヤドリギは、世界中で半寄生植物の代表例として知られています。

さらに、寄生植物には絶対寄生植物と条件的寄生植物という分類も存在します。
絶対寄生植物は、宿主がいなければ発芽さえしない、あるいは生活環を完結できない植物です。 ラフレシア や ストライガ などがこれにあたります。対して、条件的寄生植物は宿主がいなくても成長・繁殖が可能ですが、宿主がいれば寄生もするという柔軟な生き方をしています。
「菌従属栄養植物」との決定的な違い

出典:WIKIMEDIA COMMONS – Monotropastrum humile 07527(Σ64撮影)
前回の記事で紹介した「光合成をやめた植物」には、寄生植物とはまったく異なる生き方をしている仲間がいます。それが「菌従属栄養植物(きんじゅうぞくえいようしょくぶつ)」です。
両者の根本的な違いは、栄養獲得の経路にあります。寄生植物は、吸器を用いて生きた植物の維管束に直接接続し、宿主から水分と養分を奪い取ります。栄養の流れは「宿主植物 → 寄生植物」という直接的なものです。
一方、菌従属栄養植物は、菌類(特に菌根菌というキノコやカビの仲間)を仲介役として、間接的に他の植物が作った栄養を得ています。菌根菌は本来、光合成を行う植物と共生関係を結び、その植物から炭水化物をもらう代わりに、土壌から吸収した水分やミネラルを植物に提供します。
菌従属栄養植物は、この菌類に「寄生」することで、菌類が光合成植物から受け取った栄養を横取りしているのです。栄養の流れは「光合成植物 → 菌類 → 菌従属栄養植物」という複雑で間接的なものなのです。
これらは栄養獲得経路が本質的に異なりますが、進化的な共通点も見られます。どちらも光合成を放棄したことで、葉緑体ゲノムが劇的に縮小し、多くの遺伝子が失われる「還元進化」※を経験しています。
光合成をやめた植物についての概要はこちら👇️

還元進化📉
還元進化(かんげんしんか)は、生物が進化の過程で、特定の器官や遺伝子、機能などを失い、よりシンプルな体つきになることです。これは、効率を追求してエネルギーコストを節約する、積極的で合理的な「適応」戦略の一つです。
代表的な例として、光のない洞窟に住む魚(ブラインドケーブ・カラシン)が目を失う代わりに他の感覚を発達させたり、宿主の栄養に頼る寄生動物(サナダムシなど)が自らの消化管を失ったりしています。全寄生植物が光合成の遺伝子を失うのも、この還元進化の顕著な例です。

出典:WIKIMEDIA COMMONS – Astyanax jordani SeaDonuts1
微生物の世界ではさらに極端な例が見られます。アブラムシの体内に共生する細菌ブフネラ(Buchnera aphidicola)は、自由生活から共生生活へ移行する際に、遺伝子を元の約4分の1(約56万塩基対)にまで縮小させました。これにより、膨大なエネルギーを節約できますが、同時に柔軟性が失われ、宿主なしでは生きられなくなります。
人間にも還元進化の痕跡があります。尾てい骨(尾椎骨、コッキックス)や虫垂(ちゅうすい)は、かつて祖先ではより大きな機能を持っていました。クジラの骨盤も、陸上の祖先が持っていた後ろ脚の名残で、現在では生殖に関連する筋肉の付着点として限定的な機能を持つだけです。
還元進化は、不要なコストを削減できるメリットがある一方、環境変化への対応力(柔軟性)が失われるデメリットもあります。
⚠️ 「退化」との違いと用語の注意点
「退化」という言葉は、しばしば「劣るようになった」という否定的な(価値判断の)意味合いで使われます。しかし、生物の進化に「優劣」はありません。
還元進化は、環境に適応した結果として機能を失う合理的な「変化」です。そのため、科学の世界では価値判断を含まない「還元進化」や「後退進化」という言葉を使うことが推奨されます。また、「痕跡器官(vestigial organ)」は、かつて機能していたが現在は元の機能を失った器官を指し、還元進化の結果として生じたものです。
吉田 聡子, 白須 賢 ‐ ストライガのゲノム解析から見えてきた寄生植物の進化(2020年)
Biology Insights – Buchnera and Aphids: Symbiosis, Genomics, and Co-evolution
Wikipedia – Human vestigiality
The Scientist – What Does the Appendix Do?
PMC (PubMed Central) – A Glimpse Towards the Vestigiality and Fate of Human Appendix
Academia.edu – Reductive Genome Evolution In Buchnera Aphidicola
寄生植物研究の歴史 ― 古代認識からゲノム科学の最前線へ

出典:WIKIPEDIA COMMONS – Cuscuta campestris
寄生植物という存在を、人類はいつ認識したのでしょうか?古代から現代まで、この不思議な生き方をする植物たちの謎を解き明かそうとする営みは続いてきました。
古い時代の「奇妙な植物」という認識から、最新のゲノム科学による分子レベルでの理解へと至るまで、研究方法の進化が知見をどこまで深めてきたのか、その足跡をたどっていきましょう。
古代から近代への認識変化(紀元前~19世紀)
寄生植物に関する科学的な記録は、驚くべきことに古代ギリシャにまでさかのぼります。「植物学の父」と呼ばれるテオフラストス(紀元前373年頃~287年頃)は、紀元前300年ごろの著書『植物誌』の中で、すでにネナシカズラなどの特徴を記述しています。

出典:WIKIMEDIA COMMONS – Teofrasto Orto botanico detail
しかし、古代の記述は外見上の観察にとどまっていました。それらが菌類ではなく「植物である」という確実な認識は、長い間曖昧なままでした。
転機は18世紀から19世紀にもたらされます。植物学者たちの詳細な観察により、これらが花を咲かせる「植物である」ことが実証され、地中で根に寄生するタイプの存在も学術的に確立されました。この時代に、寄生植物は「奇妙な存在」から科学的な分類の対象へと変わっていったのです。
分類学と形態学の時代(19~20世紀)
19世紀後半から20世紀初頭にかけて、研究の中心は「分類」へと移りました。世界中から標本が集められ、その形態的特徴、特に花や茎、果実の形に基づいて、寄生植物の系統樹※も作られていきました。
系統樹🌳
系統樹(けいとうじゅ)とは、生き物たちの「家系図」のようなもので、進化の歴史の中で種がどのように分かれてきたのかを樹木のように表した図です。枝の分かれ目が共通の祖先にあたり、枝と枝が近いほど、生物同士が近い親戚関係にあることを示します。

出典:東京薬科大学 ‐ 動物の進化系統樹 山岸 明彦
かつては骨格や見た目の比較が中心でしたが、現在はDNA(遺伝情報)の解析が主流となり、より客観的な関係性を探れるようになりました。この系統樹は生命38億年の壮大な物語を読み解くための地図であり、新しい発見によって今もなお枝葉が更新され続けています。

出典:松井 求 ‐ 分子系統解析の最前線

出典:WIKIMEDIA COMMMONS – Phylogenetic Tree of Life-ja
ネナシカズラ属(Cuscuta)など、主要なグループの詳細な分類体系がこの時期に確立されます。その研究成果は非常に精緻なもので、現在でもその骨格は利用されています。
しかし、この時代の研究には限界もありました。収斂進化※により結果的に似た生き物の存在などもあり、外見が似ているというだけでは、必ずしも進化的に近いとは限りません。また、寄生の根本的な仕組み、つまり「なぜ」「どのように」宿主を見つけるのか、という分子レベルの問いには答えることができませんでした。
収斂進化🦉🦇
収斂進化(しゅうれんしんか)とは、まったく異なる進化の道をたどった生き物が、似たような環境に適応するため、結果的に似たような姿や能力を獲得することです。たとえば、海のイルカ🐬(哺乳類)と魚のサメ🦈(魚類)は、遠い親戚ですが、水中を速く泳ぐために流線型の体とヒレを獲得しました。
空を飛ぶコウモリ🦇(哺乳類)と鳥🕊️(鳥類)も、同じように羽を広げて飛び立ちます。これは、限られた環境で生き残るために、最も効率の良い形や能力が、偶然にも「収束」した結果なのです。

出典:Wikipedia『収斂進化』
例えば、モグラ(哺乳類)とケラ(昆虫)は、どちらも土を掘って暮らす「地中生活者」です。そのため、モグラの前脚とケラの前脚は、土をかき出すために幅広く平たい形状になった点で収斂進化しています。
分類学に興味を持った人には、こちらもおすすめ👇️

ゲノム科学による革命(21世紀~現在)
真の転機は、21世紀初頭からのゲノム解析技術(全遺伝情報の解読)の飛躍的な進展とともに訪れました。国際的な研究プロジェクトチームが、寄生の度合いが異なる様々な寄生植物の遺伝情報を網羅的に比較し始めたのです。
これにより、研究は「形」から「遺伝子」のレベルへと一気に深化します。特に、ハマウツボ科のストライガなどのゲノムが解読されたことで、寄生生活への適応の痕跡が明らかになりました。
例えば、宿主が分泌する化学物質(ストリゴラクトン※)を感知する「センサー」の遺伝子が、ゲノム内で著しく数が増えている(遺伝子ファミリーが拡大している)ことが判明しました。これは、多様な宿主を効率よく認識するための適応進化だと考えられています。
ストリゴラクトン🌿
ストリゴラクトン(Strigolactones)は、植物自身が作り出す「植物ホルモン」の一種で、植物の体内では枝の数を適切にコントロールし、姿形を整える(「頂芽優勢の制御」)という重要な役割を持っています。この機能により、植物は栄養が限られた環境で、エネルギーを効率的に配分できるのです。
しかし、ストリゴラクトンの真価は、土の中で発揮されます。根から分泌されると、土壌中に生息する「菌根菌(きんこんきん)」という有益な菌類(カビやキノコの仲間)に対して、「一緒に共生しましょう」という相利共生への「招待状」として機能します。菌根菌はこの化学シグナルを感知して、植物の根に接近し、やがて根の内部に侵入します。その見返りに、植物は水分や栄養素(特にリン)を菌から受け取ることができるのです。
出典:米山香織 ‐ 多様な骨格を持つストリゴラクトンの生合成経路(2022年)
ところが、進化の過程で、ストライガなどの寄生植物がこのシグナルを「盗み聞き(傍受)」することを学んでしまったのです。寄生植物の種子は土中で何十年も待機できますが、宿主根から分泌されたストリゴラクトンを感知した瞬間、「宿主が来た!」と感知して発芽するため、このシグナルを自らの「スイッチ」として悪用しているのです。
同じ化学物質が「共生の合図」として使われると同時に「寄生のトリガー」としても機能するという、進化における分子シグナルの多機能性を示す、極めて興味深い例です。
PMC (PubMed Central) – Strigolactones: Chemical Signals for Fungal Symbionts and Parasitic Plants

出典:WIKIMEDIA COMMONS – Striga hermonthica 1148108
驚くべき「遺伝子水平伝播」の発見
ゲノム解析は、さらに驚くべき事実を明らかにしました。それは「遺伝子水平伝播(いでんしすいへいでんぱ)」※の発見です。
通常、生物の進化において遺伝子は親から子へと「垂直」にのみ受け継がれると考えられてきました。しかし、寄生植物は吸器を通じて宿主の組織に接続する際、なんと宿主の遺伝子そのものを「水平」に取り込み、自らのゲノムの一部として機能させていたのです。
これは、他の生物の能力を文字通り「盗み取る」ことで進化するという、従来の進化の常識を揺るがす大発見でした。
遺伝子水平伝播(いでんしすいへいでんぱ)🧬
遺伝子水平伝播(いでんしすいへいでんぱ)とは、遺伝子が親から子へ「垂直」に伝わる(遺伝)のとは違い、個体間や異なる種の間で「水平」にコピー&ペーストされる現象です。つまり、遺伝子の鋳型が他の生物に直接移され、機能する、という極めて異例な進化メカニズムなのです。
特に細菌(バクテリア)の世界では日常的に行われており、細菌はこの現象を通じて、他の細菌や異なる微生物から有用な遺伝子を「借用」しています。これが、抗生物質が効かなくなる「耐性遺伝子」を仲間同士で交換し合う主な手段となり、医療現場では「スーパーバグ(超耐性菌)」が急速に蔓延する深刻な問題を引き起こしています。
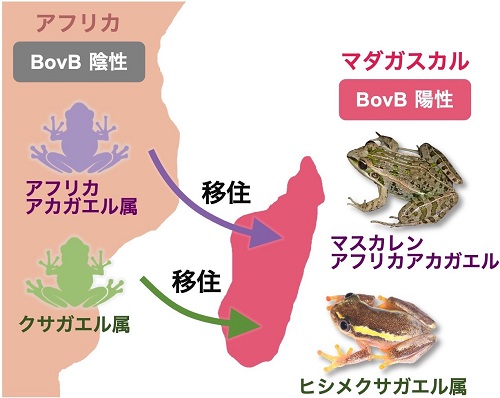
ヘビからカエルへの遺伝子水平伝播
2024年、日本の研究チームが驚くべき発見を報告しました。ヘビに特徴的な「BovB」という動く遺伝子が、被食者であるカエルに飛び移っていたのです。通常、捕食者から被食者への遺伝子移動など考えられませんが、この現象は世界中で少なくとも42回起きており、特にマダガスカルでは過去5,000万年間に14回以上も発生しています。

出典:広島大学 ‐ 【研究成果】ヘビの遺伝子がカエルに飛び移る?−寄生虫が仲介する遺伝子水平伝播のパンデミック−(2022年4月)
研究により、この仲介者はヘビとカエルの両方から吸血する寄生虫(ヒルなど)であることが判明しました。寄生虫が「遺伝子の運び屋」として機能し、異なる動物間で遺伝子を媒介しているのです。
さらに興味深いのは、アフリカからマダガスカルに移住したカエルが、移住後に初めてこの遺伝子を獲得したことです。これは人間がマラリアに感染した地域に移住する仕組みと極めて似ており、高等動物間の遺伝子水平伝播が風土病と同じメカニズムで広がる可能性を示唆しています。
出典:広島大学 ‐ 【研究成果】ヘビの遺伝子がカエルに飛び移る?−寄生虫が仲介する遺伝子水平伝播のパンデミック−(2022年4月)
細菌にとっては、突然変異を待たずに急速に進化できる強力なメリットですが、人類にとっては医療や農業上の大きな脅威となります。一方、
- 寄生植物ラフレシアが吸器を通じて宿主から遺伝子を盗む
- アブラムシが菌類から色素合成に必要な遺伝子を獲得
- 苔が水中から陸上へ進出する際に原核生物から重要な遺伝子を受け取る
など、生物の進化に大きな影響を与える現象として注目されています。
これは、進化は「親から子へ」という垂直の流れだけでなく、「種を超えた水平の流れ」によっても形作られているという、従来の進化論を書き換える発見なのです。
理化学研究所 ‐ イネ科の宿主から寄生植物へ、核内遺伝子が水平伝播する現象を発見-寄生植物が、栄養源に加えて遺伝子も宿主植物から獲得-(2010年5月)
松井 一彰 ‐ 微生物生態系における細菌の遺伝子水平伝播現象(2015年8月)
Wikipedia – Horizontal gene transfer
Nature Communications – Widespread impact of horizontal gene transfer on plant colonization of land
EBSCO Research Starters – Lateral gene transfer
PMC (PubMed Central) – Lateral gene transfer between prokaryotes and eukaryotes
PMC (PubMed Central) – Horizontal gene transfer between bacteria and animals
Royal Society Publishing – Horizontal gene transfer in evolution: facts and challenges
例えば、世界最大の花ラフレシアの研究では、働いている遺伝子の約2%が宿主から盗み出したものであったと報告されています。寄生への依存度が高い種ほど、この遺伝子の盗み取りが頻繁に起こることも分かりました。
このような植物は、宿主の能力を利用することで、自らの進化の「近道」をしていたのかもしれませんね。
社会的課題との結びつき
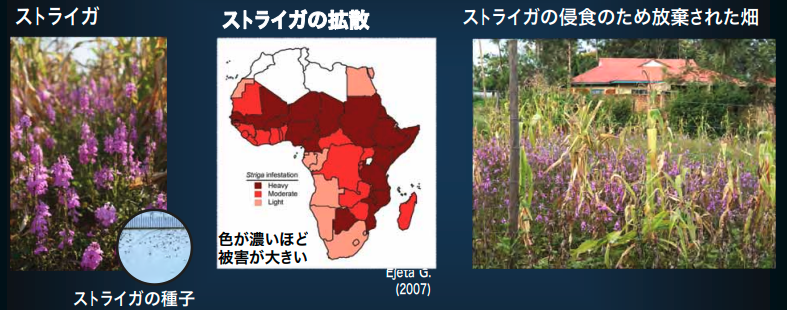
寄生植物の基礎研究は、純粋な学術的関心を超え、喫緊の社会的課題とも深く結びついています。特にアフリカのサハラ砂漠以南では、「魔女の雑草」とも呼ばれるストライガがトウモロコシやイネといった主食穀物に寄生し、甚大な食糧被害を引き起こしています。
その被害は日本の国土の1.4倍もの耕作地に及び、年間で数千億円から1兆円規模の経済損失をもたらしていると推定されています。この課題に対し、基礎研究の成果が解決策を生み出しています。
出典:内閣府 ‐ アフリカで猛威を振るう寄生植物ストライガの撲滅に向けて(2020年1月)
その代表例が「自殺的発芽」戦略です。この方法では、ストライガの種子が、宿主の化学物質(ストリゴラクトン)を感知して発芽する仕組みを逆手に取ります。
作物を植える前に、この化学物質やその類似品を畑に撒くことで、宿主がいない状態でストライガを一斉に発芽させ、寄生相手を見つけられずに枯死させるのです。
この戦略の実現性を飛躍的に高めたのが、日本の研究チームによる貢献です。2018年、彼らは「SPL7」と呼ばれる超高感度な発芽誘導物質の開発に成功しました。これは、従来の物質のわずか1万分の1という極めて低い濃度(フェムトモルレベル)でもストライガを発芽させることができます。
この発見は、「科学が社会課題を解決できる」という力強いメッセージとなりました。ごく微量で効果を発揮するため、生産コストを抑え、環境への影響も最小限にできる可能性があります。

まさに、基礎研究で寄生植物の弱点を徹底的に調べ上げたからこそ可能になった、画期的な応用研究といえますね
寄生植物研究の歴史は、寄生植物がいかに宿主と深く結びついているかを示すとともに、進化の予想外の側面を明らかにしています。基礎研究と社会課題解決が密接に結びついた寄生植物研究は、今後も人類にとって重要な学問領域であり続けるでしょう。
なぜ「寄生」という道を選んだのか? ― 進化の背景

出典:iNaturalist – Cynomorium coccineum
植物の基本的な営みである光合成。それをあえて手放すという選択は、一見すると非常に不利な道を選んだように思えるかもしれません。
しかし、生物の進化は常に合理的です。ここで紹介する植物たちが「寄生」という生き方を選んだのには、深い理由と、想像を超える革命的な戦略がありました。
光合成を維持することの「多大なコスト」と、すでにある仕組みを「侵略へ転用」した進化の巧妙さ。その二つの側面から、寄生植物がたどった進化の背景をのぞいてみましょう。
光合成のコストからの解放
私たちはつい、太陽の光さえあれば植物は無限にエネルギーを生み出せると考えがちですが、実は光合成※は非常に高コストな営みです。
光合成🌞🌿
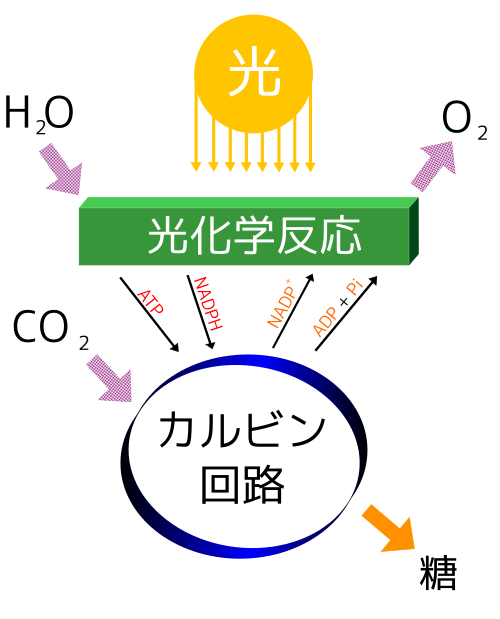
光合成は、植物や藻類(そうるい)などが、太陽の光エネルギーを利用して化学エネルギーを生み出す、地球上で最も重要な反応の一つです。太陽の光エネルギーを使い、空気中の二酸化炭素と根から吸収した水を材料にして、ブドウ糖などの有機物(栄養分)と酸素を作り出します。
このプロセスは二つの段階で進みます。まず「光反応」では、光エネルギーが水を分解して酸素を放出し、同時にATPやNADPHという形でエネルギーを蓄えます。次に「暗反応(カルビン回路)」では、そのエネルギーを使って二酸化炭素から糖を合成します。
出典:WIKIMEDIA COMMONS – Simple photosynthesis overview-ja
💡 ATP(アデノシン三リン酸)
細胞内で「エネルギーの通貨」として機能する分子です。光反応で光エネルギーが ATP に変換され、この ATP が放出するエネルギーを使って、カルビン回路で二酸化炭素から糖が合成されます。
ATP は三つのリン酸基を持ち、そのうち一つが外れるときに大量のエネルギーを放出する仕組みになっています。光合成だけでなく、あらゆる生命活動(筋肉の収縮、タンパク質合成、神経活動など)のエネルギー源として機能しており、生物学において最も重要な分子の一つです。
💡 NADPH(ニコチンアミドアデニンジヌクレオチドリン酸化物)
光反応で生成される「還元力を持つ分子」で、エネルギーと同時に電子を運ぶ役割を果たします。水が光エネルギーで分解されるとき、放出された電子がNADPHに受け渡され、NADPH はその電子をカルビン回路に運んで二酸化炭素の還元(糖への変換)を進めます。

出典:日本生物物理学会 ‐ 「光エネルギー変換を担う色素:クロロフィル」
つまり、ATPが「エネルギー」を供給し、NADPHが「還元力」を供給することで、光合成という複雑な化学反応が成立しているのです。反応式は「6CO₂ + 6H₂O + 光 → C₆H₁₂O₆ + 6O₂」で表されます。
この営みは、約27億年以上前に出現したシアノバクテリア(藍藻)という生物が始めたと考えられており、地球に酸素をもたらし、現在の多様な生命を支える食物連鎖の基盤となっています。一方で、光合成は維持するのに大量のエネルギーを必要とし、特に光が不足する環境では効率が低下します。
出典:WIKIMEDIA COMMONS – Morphological diversity in cyanobacteria
植物が光合成を行うためには、光を捉えるための葉緑素(クロロフィル)や、二酸化炭素を取り込むための酵素(こうそ)が大量に必要です。特に「RuBisCO(ルビスコ)」と呼ばれる酵素は、地球上で最も多く存在するタンパク質といわれるほどですが、これらを維持・生産するためには、莫大な量の窒素(ちっそ)が欠かせません。
窒素は、通常では土壌から吸収するしかなく、多くの植物にとって成長を左右する限られた資源なのです。
もし、他の植物が生い茂る森の暗い林床や、栄養の乏しい土地で、光も窒素も十分に得られないとしたらどうでしょう。光合成で得られるエネルギーよりも、そのための装置(葉や酵素)を維持するコストの方が大きくなってしまう、「赤字経営」に陥る可能性があります。
寄生植物は、この光合成の維持コストから自らを解放する道を選びました。光合成をやめれば、高価な「光合成工場」を維持する必要がなくなります。
その結果、暗い場所でも生きられるようになり、これまで光合成装置の維持に使っていた資源を、すべて
- 種子を多く作ること
- 宿主を探すこと
など、他の生存戦略に集中できるようになったのです。これは、厳しい生存競争を生き抜くための、非常に合理的な適応戦略でした。
吸器の起源 ― 共生から寄生への転用
寄生植物が光合成をやめることができたのは、もちろん「吸器(きゅうき)」という武器を手に入れたからです。では、この吸器はどのようにして生まれたのでしょうか。
驚くべきことに、近年のゲノム研究によって、この侵略器官はゼロから生み出された全く新しい器官ではない可能性が高いことが分かってきました。
多くの植物の根は、土壌の中にいる「菌根菌(きんこんきん)」と呼ばれる菌類(カビやキノコの仲間)と共生しています。これは、植物が陸上に進出した頃から続く古い「助け合い」の関係です。植物は菌類に光合成で作った養分を与え、菌類は土壌から集めた水分やミネラルを植物に渡します。
最新の研究によれば、寄生植物が吸器を作るために働いている遺伝子群の多くが、もともと植物が根を発達させるためや、この菌根菌と共生するために使っていた遺伝子プログラムと共通していることが分かってきました。
つまり、寄生植物の祖先は、この他者と組織的につながるための遺伝子プログラムを巧みに転用し、助け合いの相手だった菌類ではなく、「他の植物」に直接侵入し、一方的に養分を奪う「侵略の武器」へと作り変えていったと考えられています。
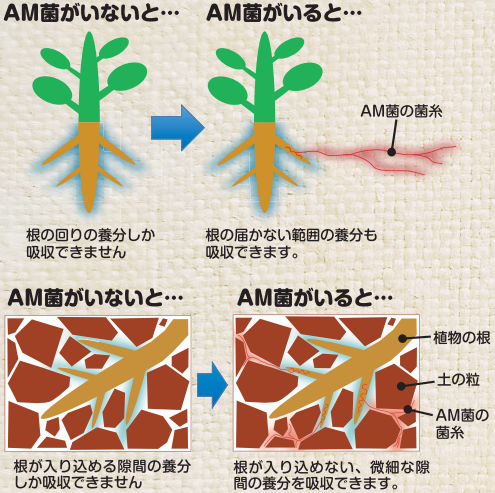

出典:農研機構 ‐ 土着菌根菌を活用することでリン酸肥料を節約できる(2014年11月)
この「助け合い」の仕組みを「侵略」の仕組みへと転換させたという進化は、植物の適応能力の柔軟さを示しています。そして、この「菌類との共生」というキーワードは、次回の記事で取り上げる、もう一つの光合成をやめた植物「菌従属栄養植物」の進化の謎を解く、最も重要な鍵ともなるのです。
寄生という生き方は、光合成の重いコストから逃れ、すでにある仕組み(根や共生のプログラム)を巧みに転用することで新たな可能性を切り開いた、きわめて賢明な進化の道筋だったといえるでしょう。
寄生植物の「戦略」 ― 宿主発見から子孫繁栄まで

出典:iNatiralist – Hydnora esculenta
※マダガスカルに生息する完全寄生植物
光合成をやめ、他者に頼るという道を選んだ寄生植物たち。しかし、彼らはただ運を天に任せているわけではありません。動くことのできない彼らは、宿主を見つけ出し、巧みに侵入し、そして子孫を繁栄させるため、精密な「戦略」を備えています。
その巧みな生存戦略を、宿主との出会いから順に見ていきましょう。
宿主を「嗅ぎ分ける」化学センサー
多くの寄生植物(特にストライガなど)の種子は、土の中で宿主が近づくのを何年も待ち続けます。その目覚めの合図となるのが、宿主の根から分泌される「ストリゴラクトン」という化学物質です。
例えば、ストライガの種子は、発芽後に自力で成長できる距離(わずか3~4mm)に宿主がいる時だけ、この物質を感知して目覚めます。これは、無駄な発芽を抑える究極の省エネ戦略です。この仕組みは「鍵と鍵穴」の関係に似ており、特定の化学物質(鍵)が種子のセンサー(鍵穴)に結合することで、発芽の合図が送られます。
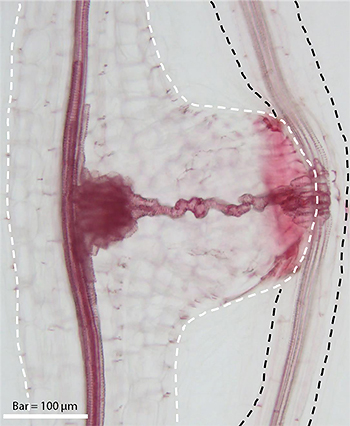
吸器 ― 宿主へ侵入する必殺の武器
宿主を認識し発芽すると、寄生植物は「吸器(きゅうき)」と呼ばれる侵入器官を形成します。吸器は宿主の組織に向かって成長し、まずは細胞の壁を溶かすための特別な酵素(β-1,4-グルカナーゼなど)を分泌します。宿主のバリアを突破すると、最終的に宿主の「維管束(いかんそく)」(水や養分が通る管)に自らの管を接続させ、栄養の通り道を完成させます。
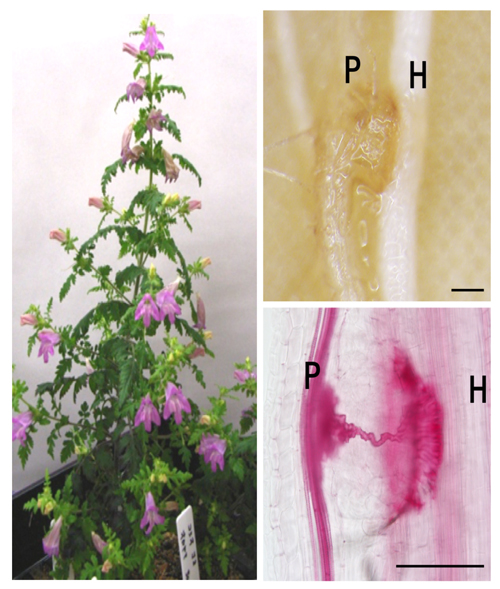
出典:基礎生物学研究所 ‐ 寄生植物の宿主植物への侵入に必要な遺伝子を同定 ~深刻な病害寄生植物の防除法開発に期待〜(2020年10月)
※左:コシオガマ植物体、右上:宿主植物(H)に寄生するコシオガマ(P)の吸器、右下:吸器内部の道管細胞を赤く染色
「接ぎ木」との共通点
この「吸器が宿主の組織と接続する」メカニズムは、私たちが農業で行う「接ぎ木」で、異なる植物の組織が癒合(ゆごう)する仕組みと非常によく似ていることが分かっています。植物が本来持つ「他者とつながる能力」を、寄生植物は一方的な「侵略」のために転用しているのです。
子孫を増やす、多様な種子散布
宿主から養分を得て成長した寄生植物は、次の世代に命をつなぐため、実に多様な方法で種子を散布します。ストライガやナンバンギセルは、埃(ちり)のように微細な「埃種子(ほこりしゅし)」を何万と作り、風に乗せて広範囲に散布します。
一方、ヤドリギは粘着性の高い果実を鳥に食べさせ、鳥がフンをしたり、くちばしを拭いたりする際に、種子を他の木の枝に「塗り付ける」戦略をとります。さらには、地中でキノコに似た姿のツチトリモチの仲間が、カマドウマ(昆虫)に種子を運ばせている例も見つかっています。

出典:京都大学 ‐ 光合成をやめたラン科植物ツチアケビにおける鳥による種子散布 -動物に種子散布を託す初めてのラン科植物の発見-(2015年5月)
※(a)ツチアケビ植物体(花期)、(b)ツチアケビ植物体(結実期)、(c)ツチアケビ花茎につかまるウグイス、(d)ツチアケビ果実を摂食するヒヨドリ、(e)ヒヨドリに摂食されたツチアケビ果実、(f)ツチアケビ果実の切片、(g)ヒヨドリ糞中のツチアケビ種子、(h)ツチアケビ種子の切片(リグニン化した種皮がサフラニンで赤く染色されている)
このように、寄生植物は宿主の「匂い」を嗅ぎ分ける化学センサー、確実に侵入するための吸器、そして次の世代を残すための巧みな種子散布戦略まで、その一生を通じて高度な生存術を発達させてきました。次の章では、こうした戦略を駆使する寄生植物には、具体的にどのような仲間たちがいるのか見ていきましょう。
寄生植物の多様な世界 ― 寄生植物の代表種とその生態

出典:iNaturalist – Pilostyles thurberi
寄生植物と一口に言っても、その姿や生き方は驚くほど多様です。緑の葉を持ちながらこっそり栄養を奪うものから、葉も茎も根も捨て去り、宿主と一体化するものまで…。ここでは、寄生植物の機能的なグループ分けに基づき、代表的な種とその独特な生態をご紹介します。
根寄生半寄生植物 ― 光合成と寄生の両立
自らも光合成を行う緑の葉を持ちながら、宿主の根から水分や養分を奪う半寄生植物。農業に甚大な被害をもたらすものから、生態系のバランスを保つものまで、その生態は多様です。
このグループには、
- 随意的半寄生植物:宿主なしでも生活できる
- 義務的半寄生植物:宿主がなければ生活環を完結できない
に分けることができ、その適応戦略の多様性が生物進化の奥深さを示しています。
ハマウツボ科 Orobanchaceae ストライガ属 Striga

出典:iNaturalist – Striga hermonthica
- 和名: ストライガ属(代表種:ストライガ・ヘルモンシカ)
- 学名: Striga (代表種:Striga hermonthica (Delile) Benth.)
- 分類: 被子植物門 双子葉類 シソ目 ハマウツボ科 ストライガ属
- 分布: アフリカ(主にサハラ砂漠以南)、アジア、オーストラリア、アラビア半島
- 種数: 約30~42種(分類上の不確実性により正確な数は不明)
- 保護状況: 該当なし(侵略的外来種として複数国で規制対象)
ストライガ属は、トウモロコシやイネ、ソルガムといった重要な穀物の根に寄生する「根寄生植物」の仲間です。宿主の栄養や水分を奪って枯らしてしまうため、英語ではその恐ろしさから「魔女の雑草(Witchweed)」と呼ばれています。

出典:神戸大学 ‐ 食糧生産に甚大な被害をもたらす寄生植物ストライガの養水分収奪機構を解明(2019年2月)
ケニアでは被害農地の60%以上でストライガが確認されており、300百万人以上の農民に影響を与えています。発芽初期は光合成色素をほぼ持たず、成熟後に光合成を始める「義務的半寄生植物」という特殊な生態が科学的に注目されています。
ハマウツボ科 Orobanchaceae イエローラトル Rhinanthus minor

出典:iNaturalist – Rhinanthus minor
- 和名: 特になし(ここでは英名のイエローラトルと呼びます)
- 学名: Rhinanthus minor
- 分類: 被子植物門 双子葉類 シソ目 ハマウツボ科 リナンサス属
- 分布: ヨーロッパ全域、シベリア、ロシア極東、中央アジア、カザフスタン、北米(導入種として北部・南西部)
- 保護状況: 該当なし(北米では雑草として認定、ただし草原再生に有用)
イエローラトル(Rhinanthus minor)はヨーロッパの牧草地などで、優占種となりやすいイネ科植物に寄生し、その成長を抑制します。この特性から、ヨーロッパでは生物多様性の高い野生草花の牧草地を復元・維持するための「ナチュラル・グラス・コントロール」として意図的に導入されており、攻撃的なイネ科植物の競争力を弱めることで、より弱小な野生花が生育できる空間を生み出しています。
かつてノルウェーなどの農業社会では、イエローラトルの種子が熟すことを「刈り入れ時期の指標」(mowing indicator)として利用していたほど、その成長パターンが広く認識されていました。英名の「Yellow Rattle(黄色いガラガラ)」は、熟した果実が風で揺れると種子がカラカラと鳴ることに由来します。
茎寄生植物 ― 空中からの侵略者
宿主の地上部(茎や枝)に寄生する植物たちが茎寄生植物です。ここでも、
- 半寄生:光合成能力を残す
- 完全寄生:完全に依存する
の二つのタイプが存在します。根寄生植物が地中で静かに栄養を盗むのに対し、茎寄生植物は宿主の地上部を劇的に変容させます。
ビャクダン科 Santalaceae セイヨウヤドリギ Viscum album

出典:iNaturalist – オウシュウヤドリギ Viscum album ssp. album
- 和名: セイヨウヤドリギ(ヤドリギ、オウシュウヤドリギ)
- 学名: Viscum album
- 分類: 被子植物門 双子葉植物綱 ビャクダン目 ビャクダン科 ヤドリギ属
- 分布: ヨーロッパ、北アフリカ、西・中央アジア
- 習性: 半寄生常緑低木
- 保護状況: 低懸念(LC)
セイヨウヤドリギ(Viscum album)は、他の樹木の枝や幹に吸器(特殊化した根)を差し込む「半寄生植物」の代表例です。緑色の葉で自らも光合成を行いますが、宿主の木から主に水分やミネラル(無機栄養)を奪って生活しています。
200種以上の樹木に寄生可能で、リンゴ、ポプラ、ボダイジュ、モミ、マツ、カラマツなど、宿主によって異なる亜種が認識されています。英語では「mistletoe(ミスルトウ)」と呼ばれ、ヨーロッパでは古くから神聖な植物とされてきました。古代ケルト人のドルイド僧は冬至の祭りで神聖視し、現代ではクリスマスの飾りとして世界的に有名です。

出典:iNaturalist – ヤドリギツグミ Turdus viscivorus
粘着性の高い白色または黄色の果実を、特にヤドリギツグミ(Turdus viscivorus)などの鳥に食べさせ、鳥が糞とともに、あるいはくちばしをこすり付けることで、種子を他の木の枝に運んでもらうという巧みな散布戦略を持っています。気候変動の影響を受けやすく、現在ヨーロッパでは分布域が北東方向へシフトしている ことが報告されています。
ヒルガオ科 Convolvulaceae アメリカネナシカズラ Cuscuta campestris

出典:iNaturalist – アメリカネナシカズラ Cuscuta campestris
- 和名: アメリカネナシカズラ(アメリカ根無葛)
- 学名: Cuscuta campestris Yunck.
- 分類: 被子植物門 双子葉植物綱 ナス目 ヒルガオ科 ネナシカズラ属
- 分布: 北米原産、現在は北米、中米、南米、ユーロ・メディテラネア地域、シベリア、中央アジア、西アジア、インド、アジア全域、アフリカ、オーストラリアに定着
- 習性: 全寄生一年生つる植物
- 保護状況: 侵略的外来種(多くの国で規制対象)
アメリカネナシカズラ (Cuscuta campestris)は、北米原産の全寄生つる植物で、クローバーを模倣する細い黄緑色のつるを持ちます。アルファルファ(ルーサン)、トマト、ニンジン、砂糖大根、その他200種類以上の植物に寄生でき、吸盤状の吸器を宿主に貫通させ、養分と水分を直接略奪します。
光合成は殆どできず、完全に宿主に依存する「全寄生植物」で、地球上で最も広がった侵略的雑草として、ヨーロッパ、アジア、アフリカ、オーストラリアなど世界中で問題になっています。

出典:iNaturalist – アメリカネナシカズラ Cuscuta campestris
特に農業地帯では深刻な被害をもたらし、1株のアメリカネナシカズラが最大16,000粒の種子を生成でき、わずか2ヶ月で5メートル成長して周囲の植物を完全に被覆することもあります。一方で、中国南部ではこのアメリカネナシカズラが、侵略的な外来種 ホシアサガオ(Mikania micrantha)の制御に自然の「天敵」として機能する可能性が報告されており、同じ侵略者が、別の侵略者を抑制するという複雑な生態系が浮かび上がります。
ニュージーランドなど、その他多くの国では、国立公園や湿地保護区での撲滅に積極的に取り組んでいます。
根寄生完全寄生植物 ― 地中に潜むスペシャリスト
根寄生完全寄生植物は、光合成能力を完全に失い、生活のすべてを宿主の根に依存する植物たちです。地上には花だけを咲かせるものが多く、その生態は長く謎に満ちていました。
これらの植物は、寄生生活への進化が極端に進んだ、生物多様性の中でも最も個性的な存在です。
ハマウツボ科 Orobanchaceae ハマウツボ属 Orobanche

出典:iNaturalist – Orobanche cumana
- 和名: ハマウツボ属
- 学名: Orobanche
- 分類: 被子植物門 双子葉植物綱 シソ目 ハマウツボ科 ハマウツボ属
- 分布: 温帯北半球(主にユーラシア大陸)、北アフリカ、亜熱帯・熱帯アフリカ、オーストラリア、ニュージーランド
- 種数: 約170~200種
- 習性: 全寄生一年生または多年生草本植物
- 保護状況: 侵略的外来種・検疫対象(多くの国で農業害虫として規制)
ハマウツボ属(Orobanche )は葉緑素を持たず光合成できない根寄生完全寄生植物です。ヨモギ、トマト、タバコ、マメ科作物などに寄生する種も多く、産業の場面では世界的に深刻な農業被害をもたらす有害雑草として知られています。

出典:iNaturalist – ヤセウツボ Orobanche minor
ハマウツボ属は世界で約1,600万ヘクタールの農地に広がり、作物の20~100%を枯死させることで、1億人以上の生計に影響を及ぼしていると推定されています。1株が10,000~1,000,000粒の微細な種子(埃種子)を生産し、風で長距離散布されるため、一度侵入すると根絶が極めて困難です。
同時に、宿主依存性・種子休眠・発芽誘導物質(ストリゴラクトン類)との関係が研究対象としても注目されています。寄生植物は宿主の防御分子を「宿主認識の合図」として進化的に転用しており、植物の防御システムそのものが寄生のトリガーとなっているのです。
ハマウツボ科 Orobanchaceae ナンバンギセル Aeginetia indica

出典:iNaturalist – ナンバンギセル Aeginetia indica
- 和名: ナンバンギセル(南蛮煙管)
- 学名: Aeginetia indica L.
- 分類: 被子植物門 双子葉植物綱 シソ目 ハマウツボ科 ナンバンギセル属
- 分布: 熱帯・亜熱帯アジア(インド、中国、日本、東南アジア)、ニューギニア
- 習性: 全寄生一年生草本植物
- 保護状況: 低懸念(LC)、ただしスリランカでは「絶滅危惧IA類/絶滅の可能性」
ナンバンギセル(Aeginetia indica)は、葉緑素を全く持たない根寄生完全寄生植物です。ススキやサトウキビなどイネ科植物の根に寄生し、養分と水分を奪います。
和名の由来は、赤紫色の花が南蛮(なんばん)渡来のタバコのパイプ(煙管 キセル)です。日本では古くから知られ、ネパールでは祭祀(さいし)にも使われますが、サトウキビ畑などでは農業被害をもたらす有害雑草でもあります。
最新のゲノム研究により、光合成の遺伝子を失うと同時に、宿主から84個もの遺伝子を「遺伝子水平伝播」によって獲得していたことが判明しました。
エンドパラサイト ― 宿主と一体化する究極の寄生
エンドパラサイトは寄生植物の中でも最も特異な進化を遂げたグループです。葉も茎も根も持たず、自らの体を宿主の組織内部に菌糸のように埋め込み、花だけを外部に咲かせる究極の「内部寄生植物」です。これらの植物の存在は、生命がいかに極端な環境に適応できるかを示す、生物進化の驚異的な例といえます。

「エンドパラサイト」は、寄生生物が宿主の組織内部(根の内部や幹の内部)に完全に埋め込まれている状態を指します。外部寄生(エクトパラサイト)のように宿主の表面に付着するのではなく、宿主の細胞内や組織間隙に菌糸状の本体を形成し、そこから栄養を吸収します。
ラフレシア科 Rafflesiaceae ラフレシア Rafflesia arnoldii

出典:WIKIMEDIA COMMONS – Rafflesia arnoldi 2013-12-31 21-48
- 和名: ラフレシア(世界最大の花)
- 学名: Rafflesia arnoldii
- 分類: 被子植物門 双子葉植物綱 キントラノオ目 ラフレシア科 ラフレシア属
- 分布: スマトラ、ボルネオ(インドネシア、マレーシア)の熱帯雨林
- 習性: 内部全寄生一年生(多年生サイクル)
- 保護状況: 絶滅危惧(VU:脆弱)
十数種確認されているラフレシア属(Rafflesia)のなかでも、ラフレシア(ラフレシア・アルノルディイ Rafflesia arnoldii )は、世界最大の単一花として知られています。2020年1月にインドネシア・西スマトラで発見された個体は直径111cm、重量11kg以上に達しました。
その腐肉を思わせる強い臭いから「corpse flower(腐肉花)」と呼ばれ、ハエなどの送粉者を誘引します。ブドウ科のTetrastigmaというつる植物にのみ寄生するエンドパラサイトで、本体は宿主の組織内に糸のように埋め込まれており、花のみが地表に現れます。
更に注目すべき特徴は、核ゲノムの2~3%、ミトコンドリアの最大50%が宿主由来という遺伝子水平伝播で、宿主の免疫から逃れる「遺伝的カモフラージュ」ではないかと考えられています。2023年の調査では全42種が絶滅危惧種に分類されるべきと推奨され、生育地の67%が保護区外にあることから、保全が急務です。
ヤッコソウ科 Mitrastemonaceae ヤッコソウ属 Mitrastemon ― 根から顔を出す「ヤッコさん」

出典:iNaturalist – Mitrastemon yamamotoi var. kanehirai
- 和名: ヤッコソウ属
- 学名: Mitrastemon (代表種 Mitrastemon yamamotoi )
- 分類: 被子植物門 双子葉植物綱 ツツジ目 ヤッコソウ科 ヤッコソウ属
- 分布: 日本(四国・九州)、台湾、東南アジア(M. yamamotoi)、メキシコ・中米(M. matudae)
- 種数: わずか2種(世界でも極めて稀少な科)
- 習性: 根内部全寄生多年生草本植物
- 保護状況: 日本各地で天然記念物指定、絶滅危惧
ヤッコソウ属(Mitrastemon )は、世界でもわずか2種からなる、分類学的にも希少なヤッコソウ科の植物です。ヤッコソウはエンドパラサイトで、葉緑素を持たず、ブナ科樹木の根に内部寄生し花だけを地表に出します。
日本に生息するヤッコソウ(M. yamamotoi )は1909年に牧野富太郎博士※が命名し、花の姿が大名行列の「奴(やっこさん)」に似ることから名付けられました。四国・九州を中心に分布し、各地で天然記念物として保護されています。
1909年に牧野博士が学術的に記載した Mitrastema yamamotoi の学名は、1907年にこの植物を発見した高知師範学校教諭・山本一先生への献名です。
花は雌雄同株で、段階的に性別を変えるという特殊な性戦略を持ち、最初に雄花として花粉を放出し、その後に雌花へと転換します。近年の研究では、ミトコンドリアゲノムにおいて宿主由来遺伝子を水平伝播(遺伝子水平伝播)で獲得していることが明らかになり、寄生植物の進化研究に重要な知見を与えています。
牧野富太郎(理学博士、植物学者)🌿🌸🔬
牧野富太郎博士は、「日本の植物分類学の父」と呼ばれ、日本を代表する植物学者(1862~1957年)です。高知県佐川町の酒造商の家に生まれ、小学校を中退しながら独学で植物学を究め、生涯をかけて日本全国の植物標本を40万点以上も収集しました。

出典:WIKIMRDIA COMMONS – Dr. Makino Tomitaro
牧野博士の特に大きな功績は、これらの植物に科学的な分類体系を初めて本格的にもたらしたことにあります。約1,500種類以上に学名を付け、その中の約1,350の学名が現在も国際的に使用されているという、極めて顕著な国際的貢献を果たしました。
また、牧野博士が描いた精密な「牧野式植物図」は、1,700種類以上に及び、科学的な正確さと芸術性を兼ね備えた第一級の学術資料として世界的に高く評価されています。博士の集大成である『牧野日本植物図鑑』は、1940年の発行から85年以上経った今もなお、植物同定と分類学の基礎文献として世界中の研究に活用されており、膨大な標本は「牧野標本館」(現・東京都立大学)で保管・提供され続けています。

牧野富太郎博士はなんと文久2年(1862年)5月22日生まれ、つまり明治時代が始まる6年前の「江戸時代生まれ」です!
彼の業績は、開国後の日本において、国際的な科学コミュニティに対して行った極めて重要な貢献として認識されており、毎年5月22日は「植物学の日」として制定され、博士の誕生日と功績を祝っています。
出典:高知県立牧野植物園 ‐ 牧野富太郎物語
Makino Botanical Garden Official Site – Dr. Tomitaro Makino (1862~1957)
Nebg.org – Tomitaro Makino (1862 – 1957)
Tokyo Metropolitan University Makino Herbarium Fund
Tokyo Metropolitan University Herbarium Details – Makino Herbarium
寄生植物の最新研究 ― 解明される謎と新発見

出典:iNaturalist – Misodendrum linearifolium
寄生植物研究の最前線では、次々と驚きの発見が報告されています。長らく謎とされてきた寄生植物の生存戦略が、分子レベルで解明され始めており、農業被害の軽減や生態系の理解に向けた新たな可能性が広がっています。
なぜ「自分」に寄生しないのか? ― 自己回避メカニズムの発見(2025年)
寄生植物は次々と周囲の植物に取り付いていくのに、なぜ自分自身や身近な仲間には寄生しないのでしょうか?
2025年10月、奈良先端科学技術大学院大学の吉田聡子教授らの研究チームは、この長年の謎を分子レベルで初めて解き明かしました。研究チームが調べたコシオガマという寄生植物には、「UGT72B1」という酵素が存在し、この酵素が「自分たちが出すシグナル」に糖(グルコース)をくっつけることで、そのシグナルを無力化していることが分かったのです。
寄生植物が宿主を見つける際、目印にするのが宿主の根から分泌される特定の化学物質(吸器誘導物質)です。寄生植物は、このシグナルを感知すると吸器を形成し、寄生を開始します。
しかし、寄生植物自身やその近縁種も、宿主が出すものと非常によく似たシグナル分子を分泌しています。もし、この「自分(仲間)が出すシグナル」にも反応してしまうと、自分自身や仲間同士で寄生しあうという非効率な事態(自家寄生)が起こり得ます。
コシオガマは、「UGT72B1」という特定の酵素(配糖化酵素)で、自分自身のシグナル分子に糖(グルコース)を結合させることで、シグナル分子の化学構造を変化させ、センサーが認識できない形に「不活性化」します。つまり、自分が出すシグナルを自ら隠しているのです。
実際に、この「UGT72B1」酵素が機能しなくなったコシオガマ(spoh1変異体)では、自己回避ができなくなり、自分自身のシグナルに反応して自分の根に吸器を形成しようとする現象が確認されています。
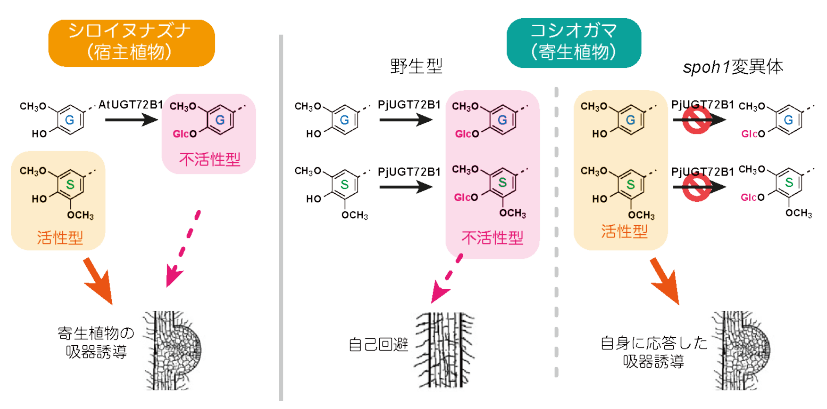
出典:京都大学 ‐ 寄生植物は自分の仲間をなぜ襲わない?~「自己回避」の仕組みを世界で初めて解明~寄生雑草の新たな防除戦略に期待(2025年10月)
農業分野では、この発見を逆転の発想で利用し、作物にこの「ブレーキ酵素」を持たせることで、寄生植物に「同じ仲間だ」と誤認させて攻撃を回避する新しい防除方法の開発が期待されています。
発芽シグナル(SL)を「自作」する能力とその本当の目的(2025年)
ストリゴラクトン(SL)は、もともと普通の植物が「枝分かれを抑える」などのために使う、自分用の「植物ホルモン」です。寄生植物は、宿主(他の植物)が出すストリゴラクトンを「発芽の鍵(合図)」として利用します。そのため、自らストリゴラクトンを作る能力は、進化の過程で失った(還元進化した)と長らく考えられてきました。
しかし2025年、明治大学や神戸大学などの国際共同研究チームは、ヤセウツボやコシオガマがストリゴラクトンを自ら生産する能力を保持していたことを発見しました。
研究チームの見解によれば、この能力は、ストリゴラクトンを発芽した後の自分自身の成長(枝分かれなど)を調節するために維持されていると考えられます。宿主の合図(SL)を悪用しつつも、自分の成長を制御する自律的な能力(SL生産)は捨てていなかったという、彼らの高度な適応戦略を示す発見です。
宿主も黙ってはいない! ― トマトの防衛センサー「CuRe1」(2020年~)
一方、被害を受ける宿主側の植物も、ただ黙ってやられているわけではありません。2020年、ドイツの研究チームは、トマトが寄生植物を「敵」として認識する仕組みを発見しました。トマトの細胞表面には「CuRe1」という受容体があり、これがネナシカズラという寄生植物の細胞壁に含まれる特定のタンパク質を察知するのです。
このCuRe1が危険をキャッチすると、トマトは直ちに防御遺伝子の発現スイッチを入れて、全身の免疫態勢に入ります。このような寄生植物と宿主との間の「攻撃(寄生植物の侵入)と防御(宿主の免疫応答)」という分子レベルでのせめぎ合いが解明されたことで、将来的には「CRISPR」などの遺伝子編集技術を用いて、特定の寄生植物に抵抗力を持つ作物品種の開発が期待されています。
このように、最新科学は寄生植物のイメージを次々と塗り替えています。彼らは単なる略奪者ではなく、環境に適応するために極めて高度な遺伝的戦略を進化させた、驚くべき生命体なのです。これらの研究成果は、寄生植物の巧妙な生存戦略を分子レベルで明らかにするとともに、農業や生態系保全への実用的な応用可能性を示しています。
生命の「つながり」を見つめ直す ― 寄生植物が教えてくれること

出典:iNaturalist – Hydnora visseri
世界最大の花ラフレシアから、農業を脅かすストライガまで、光合成をやめて他者から奪う「寄生植物」の驚異的な生存戦略…。これらの植物は決して「怠け者」などではなく、環境に適応するために極めて高度に進化した、たくましい生命の姿です。
この生き様は「チーター(cheater)」に見えるかもしれません。しかし自然界というゲームの盤上で、唯一のルールは「生き残り、子孫を残すこと」、つまり自然選択です。
寄生植物の「吸器」や「光合成の放棄」という能力は、進化というプロセスが数百万年かけて精密に打ち出した、正規の戦略であり極限の適応なのです。
「知る」ことが「守る」ことにつながる
一見、私たちの生活とは無関係に思えるかもしれない、寄生植物の生態研究…。しかし、こうした「基礎研究」こそが、現実の課題を解決する力になります。
例えば、ストライガがなぜ深刻な農業被害をもたらすのか、その弱点(発芽の仕組みや宿主認識の仕組みなど)はどこにあるのか?など、その生態を基礎研究の積み重ねにより分子レベルで徹底的に知ることで初めて、「誘引剤で発芽を誘導し、宿主がない環境で枯死させる」といった賢明な対策を生み出し、食糧という暮らしの基盤を守ることにつながっていくのです。
これは、基礎科学が社会課題の解決には欠かせないことがわかる、重要な事例の一つです。
基礎研究という土台あってこその応用研究なのです
光合成をやめた植物たちの多様な選択
宿主の遺伝子さえも取り込むラフレシアの姿は、生命が他者とつながり、影響を与え合う存在であることを示しています。
しかし、光合成をやめた植物の物語は、これだけではありません。宿主の植物から「直接」栄養を奪う寄生植物に対し、菌類(キノコやカビ)を仲介役として、森のネットワークから「間接的」に栄養を得ようとする、もう一つの不思議な仲間たちもいます。
生命のつながり方の多様さは、「直接的な略奪」にとどまらず、「共生」や「仲介」といった、より複雑で繊細な関係性をも生み出してきたのです。
次回の記事では、菌類との共生によって栄養を得る「菌従属栄養植物」たちの、奥深い世界を探っていきます。
光合成を失った彼らが、どのようにして生き続けるのか、その謎と新発見の数々をお楽しみに…!
菌従属栄養植物についてもっと詳しく知りたい人は、こちら👇️

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

【広告】この記事は「SWELL」というWordPressテーマで執筆しています。有料ですが買い切りなので、長く快適にブログを続けたい人にはおすすめですよ!👇️
参考・引用
「寄生植物」とは何か? ― 宿主から直接奪う生き方
NAIST Edge BIO – 寄生植物は宿主にどう入り込むのか?
RIKEN Research News – The genome and transcriptome of the parasitic plant Striga have been sequenced
NAIST Plant Symbisis Laboratory – Research | What Is a Parasitic Plant?
Frontiers in Plant Science – Molecular dissection of haustorium development in parasitic plants
Phys.org – Cell wall degrading enzyme is integral for plant parasitism and cross-species grafting
Virginia Tech – Westwood Lab – Parasitic plants · Plant communication · Weed evolution
AATF Africa – Fostering food security in smallholder farms in Striga weed hotspots of Kenya
寄生植物研究の歴史 ― 古代認識からゲノム科学の最前線へ
PMC (PubMed Central) – The plant vampire diaries: a historic perspective on Cuscuta research
Wikipedia – Historia Plantarum (Theophrastus)
University of Northern Iowa – Notes on Parasitic Seed Plants with Reference to Cuscuta
NAIST (Nara Institute of Science and Technology) – The genome of the parasitic plant Striga
RIKEN Press Release – 病害寄生雑草ストライガの全ゲノム解読に成功
BMC Genomics (PubMed) – Horizontal transfer of expressed genes in a parasitic flowering plant
Pest Management Science (PubMed) – Suicidal germination for parasitic weed control
なぜ「寄生」という道を選んだのか? ― 進化の背景
New Phytologist – A journey through signaling in arbuscular mycorrhizal symbiosis
Science – Plant evolution driven by interactions with symbiotic and pathogenic microbes
PMC (PubMed Central) – A roadmap of haustorium morphogenesis in parasitic plants
寄生植物の「戦略」 ― 宿主発見から子孫繁栄まで
Wikipedia – Parasitic plant: Seed germination
Phys.org – Cell wall degrading enzyme is integral for plant parasitism and cross-species grafting
PMC (PubMed Central) – Molecular dissection of haustorium development in parasitic plants
Kobe University – Plants offer fruit to insects to disperse dust-like seeds
Awkward Botany – Camel Crickets and the Dust Seeds of Parasitic Plants
Kobe University – Parasitic plants rely on unusual method to spread their seeds
寄生植物の多様な世界 ― 寄生植物の代表種とその生態
Woomer et al. (2008) – Striga management and the African farmer
AATF Africa (2021) – Fostering food security in smallholder farms in Striga weed hotspots of Kenya
PMC (2025) – Application of bioinoculants in combating Striga Hermonthica in Maize Production
Goret et al. (2021) – A decision-making tool for restoring lowland grasslands in Europe
Briggs (2021) – Mistletoe, Viscum album (Santalaceae), in Britain and Ireland
Walas et al. (2022) – The future of Viscum album L. in Europe will be shaped by climate change
Krasylenko et al. (2020) – The European mistletoe (Viscum album L.): distribution and host range
CABI Digital Library – Cuscuta campestris (field dodder)
Zagorchev et al. (2025) – Methods of Control of Parasitic Weeds of the Genus Cuscuta
Delavault et al. (2015) – Knowing the Parasite: Biology and Genetics of Orobanche
Chen et al. (2020) – The loss of photosynthesis pathway and genomic locations
Kado & Innan (2018) – Horizontal Gene Transfer in Five Parasite Plant Species in Orobanchaceae
Guinness World Records – Largest single flower
Graf et al. (2018) – Footprints of parasitism in the genome of Rafflesia lagascae
Museum of Modern Art Tokushima – Guidebook
Suetsugu et al. (2021) – Parasitic plant Mitrastemon yamamotoi: Ecology and conservation
寄生植物の最新研究 ― 解明される謎と新発見
奈良先端科学技術大学院大学 – 寄生植物が自分自身に寄生しない仕組みを解明(2025年)
PubMed – Divergent Receptors Shape Strigolactone Perception in a Facultative Parasitic Plant
PMC – CRISPR gene editing to improve crop resistance to parasitic plants
Crop Science Centre – CRISPR: The cutting-edge solution to tackle parasitic plant threats
コメント